横浜市立大学
理学部 理学科
国際総合科学部 生命環境コース
大学院 生命ナノシステム科学研究科 生命環境システム科学専攻
植物発生生理学研究室
研究紹介
詳しい研究内容
ニンジン種子の発達過程における、生重量・ABA含量・水分含有率を測定した結果、種子の成熟後期にABA含量の一時的な増加が見られ、その後種子中の水分が減少することが確認されました。この時、種子中の受精胚は魚雷型胚まで成長しており、形態的な発達は終了していました。このように、種子では乾燥が始まる前にABAが増加し、そのABAによって乾燥耐性が獲得されていると考えられます。
現在は受精胚を用いた研究は行っていません。不定胚をモデルとして、生理学的・生化学的・分子生物学的手法で研究を行っています。
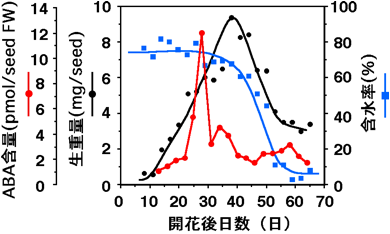
(図)ニンジン種子発達時のABA含量、生重量、含水率の変化
種子の乾燥耐性や休眠性は、ABAによって調節されています。一方、不定胚にはABAがほとんど存在しません。そのため、乾燥耐性はなく乾燥させると死滅します。また、休眠も起こらないため、続けて培養すると発芽してめばえに成長します。
ニンジン不定胚に外部からABAを処理すると、乾燥耐性と休眠性が誘導されました。乾燥耐性は、ABAの処理濃度(0.1 mMまで)や処理時間(10日間まで)を増加させるに従って高くなりました。
このように、不定胚ではABA処理(濃度、時間、発達時期など)を操作することで、乾燥耐性や休眠性を人為的に調節できることが可能です。
7-10日間のABA処理は一般的な処理時間より長いですが、これは乾燥耐性に必要な物質(タンパク質や適合溶質など)を合成し蓄積するのに必要な時間と考えられます。また、若い胚(球状胚や心臓型胚)ではABA処理を行っても乾燥耐性と休眠性は誘導されません。球状胚や心臓型胚はABAが存在しても発達を続け、魚雷型胚にまで成長します。胚の発達段階に関わる何らかの因子が、ABAの反応を調節していると考えられます。
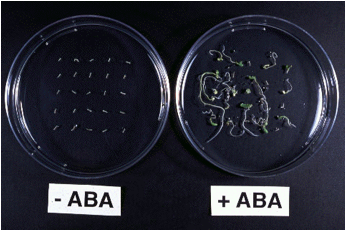
(図)乾燥処理後2週間培養した不定胚(左:ABA未処理、右:ABA処理)
ニンジンでは、乾燥した種子、乾燥した不定胚とも含水率は5%程度と、ほぼ完全に水分を失った状態にありました。つまり種子や胚の乾燥耐性は、水分を保持することではなく、水分を失っても生存できる状態をつくることにあります。水分が存在しないと細胞内の代謝が低くなるため、細胞は休眠状態になると考えられます。
また、一般に細胞は超低温にさらされると、細胞内の水分が凍結し、生じた氷の結晶によって細胞が破壊されると考えられています。乾燥した不定胚では凍結する水分が存在しないため、氷点下でも生き延びることができます。
実際に、乾燥したニンジン不定胚は超低温(-80度)で3年以上生き延びることが確認できました。この現象は、細胞をより自然に近い状態(種子休眠)で長期間保存する技術として、応用できる可能性があります。


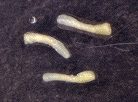
(図)ニンジン不定胚の乾燥
(左:乾燥前、中:3時間の乾燥後、右:再吸水後)
一般に植物体にABAを処理すると、多くの遺伝子が発現します。種子発達の後期においても、多くの遺伝子がABAによって発現誘導されます。
これらの代表的なものが、Late embryogenesis abundant protein(LEAタンパク質)をコードする遺伝子群です。LEAタンパク質は親水性の高いアミノ酸を多く含むことから、タンパク質分子中に水分子を保持し、細胞から水が失われるのを防ぐはたらきがあると考えられています。
他にも、アミノ酸の合成酵素や糖代謝酵素などの発現が見られます。これらは、アミノ酸や多糖類、糖アルコールなどの合成に関与します。このような物質には、細胞の浸透圧を調節する、水分子の代役としてはたらく、などの機能が予想されています。これらの遺伝子産物が乾燥耐性や休眠性にどのように関与しているのか?、どのようなメカニズムで細胞が乾燥耐性を獲得するのか?、詳細については不明です。
ニンジン不定胚からも、ABA処理で誘導される遺伝子が単離されました。その中には、LEAタンパク質や糖代謝酵素をコードする遺伝子が含まれていました。これらの遺伝子は、不定胚だけでなく種子成熟後期でも発現が見られました。さらにこれらの遺伝子は、種子や胚でのみ発現するタイプ(ECP31,ECP40,ECP63,CAISE3)、葉でも発現するタイプ(CAISE1,CAISE4,CAISE5)に分類することができます。現在は、これらの遺伝子の機能や発現調節機構を明らかにするため、詳細な発現解析や形質転換植物の解析を行っています。
遺伝子の発現調節は、転写調節因子というタンパク質のはたらきによって行われています。種子でのABAの情報伝達に関わる転写調節因子としては、VP1/ABI3因子が知られています。このVP1/ABI3因子がABAで誘導される遺伝子の発現調節を行っています。
ニンジンにおいても、その類似遺伝子としてC-ABI3が単離されました。C-ABI3を過剰発現させた形質転換植物の研究から、上で示したニンジンのABA誘導性遺伝子(種子や胚でのみ発現するタイプ)の発現が、C-ABI3とABAによって調節されていることが明らかになりました。さらに、培養細胞でC-ABI3を過剰発現させた場合には、ABA処理によって複数の遺伝子発現が誘導されるようになり、その結果乾燥耐性が獲得されました。
これらのことから、体細胞不定胚においても、VP1/ABI3因子によるABAの情報伝達、ABA誘導性遺伝子の発現、乾燥耐性の獲得など、ABAによる一連の反応が受精胚と同じように起きることが明らかになりました。
現在は、不定胚の特徴(同調的かつ大量に調製できる)を生かし、C-ABI3と関連して機能する転写因子(群)について検出を試みています。
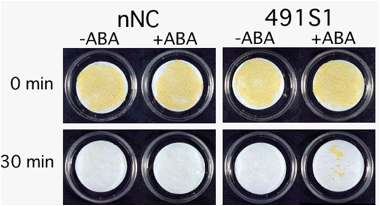
(図)乾燥処理(0分間あるいは30分間)後、3週間培養したニンジン培養細胞
nNC:遺伝子導入していない培養細胞、491S1:C-ABI3を過剰発現させた培養細胞、
それぞれABA処理をしたもの(+ABA)、しないもの(-ABA)
ニンジン不定胚やトマト種子を使って、胚発生で発現し機能するアクアポリンについて、遺伝子レベルでの解析を行っています。
細胞の内外の水分子の移動は、細胞膜の半透性によって起きると考えられてきました。しかし最近になって、細胞膜に存在する膜タンパク質(アクアポリン)が選択的に水分子を通過させている事実が示されました。水チャネルは広く生物界に保存されており、種子植物でも30種類以上のアクアポリンが存在すると考えられています。胚発生でも、胚の発生・成長や休眠・乾燥などに関わる水チャネルが存在すると考えられます。
トマト種子では、少なくとも6種類の原形質膜アクアポリン(PIP)遺伝子が発現していました。種子でのPIP遺伝子の発現は、葉での発現に比べると弱いものでしたが、6種類のPIP遺伝子はいずれも種子発達の前半で発現が強く、後半になると弱くなるという共通性が見られました。PIP遺伝子の発現が強い種子発達前期は、トマト種子が水分含有率を減少させる時期と一致していました。これらのことから、トマト種子の水分減少に原形質アクアポリンが関与している可能性が示唆されました。
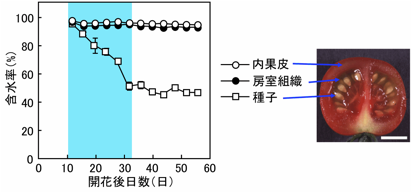
(図)トマト果実と種子の含水率の変化
水色は種子の含水率の変化が大きい期間
ニンジン不定胚からは、4種類の原形質膜アクアポリン(PIP)遺伝子と1種類の液胞膜アクアポリン(TIP)が単離されました。これらのニンジンPIP遺伝子とTIP遺伝子が、胚や種子の発達にどのように関与しているのかを、発現や機能を解析して研究を進めています。
テッポウユリ花粉プロトプラストからを利用したアクアポリンの機能解析法を開発しました。テッポウユリ花粉に由来するプロトプラストには、大型(直径0.1 mm)、均質、大量に得られるという特徴があります。さらに、原形質膜アクアポリンが発現していないこと、遺伝子導入が可能なことから、アクアポリンの機能解析に適している。テッポウユリ花粉プロトプラストにアクアポリン遺伝子を発現させて、低張液に移します。低張液中では、プロトプラスト内に水が流入しますが、この時にアクアポリンが存在すると、水の流入が速くなり、プロトプラストの膨張も速くなります。

(図)アクアポリン遺伝子を導入したテッポウユリ花粉プロトプラスト
等張液中(左)、低張液中(中)、レポーターの蛍光による遺伝子導入の確認(右)
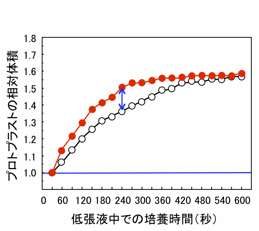
(図)低張液中での花粉プロトプラストの体積変化
アクアポリンを導入した場合(●)は、導入していない場合(○)に比べて体積の増加が速い
海で生活する種子植物(アマモ)は、種子植物であるため種子をつくります。陸上の種子植物で明らかになった知見を活かして、アマモの種子の発達と発芽についても研究を進めています。また、研究活動に関連して、海の環境再生活動にも参加しています。


(図)アマモの植物体(左)、種子(右)
2019年4月1日更新